





L’analisi metabolomica del latte rivela le pirimidine come possibili marcatori discriminanti di diverse strategie nutrizionali basate sull’utilizzo dell’insilato di mais.
Nel Nord Italia, l’insilato di mais rappresenta fino al 90% della componente foraggera nella dieta delle vacche in lattazione, a causa della fertilità del suolo, del clima favorevole e dell’elevata resa in sostanza secca per ettaro (Borreani et al., 2013; Bellingeri et al., 2019). Di conseguenza, la maggior parte degli allevamenti di quest’area geografica ha adottato strategie nutrizionali basate sulla somministrazione TMR (total mixed ration), in cui l’insilato di mais è l’ingrediente principale della razione, in combinazione o meno con altri insilati di erba, leguminose o graminacee.
Lo studio della nutrizione animale può in generale fornire informazioni importanti all’interno delle aziende lattiero-casearie, in grado di guidare i successivi processi di caseificazione, approfondendo lo studio dei cambiamenti chimici del latte in accordo al tipo di strategia nutrizionale e successiva caseificazione. Negli ultimi anni, tecniche analitiche all’avanguardia come l’analisi metabolomica hanno permesso di approfondire quella che è la composizione del latte in accordo a numerosi fattori esterni ed interni all’impianto di trasformazione, focalizzandosi in particolare sui benefici sanitari ed economici (Zhu et al., 2021). La metabolomica rappresenta lo studio di molecole organiche a basso peso molecolare (< 2.000 Dalton) coinvolti in diversi processi metabolici. I possono essere misurati con tecniche su larga scala e ad alto rendimento, tra cui la risonanza magnetica nucleare e la spettrometria di massa ad alta risoluzione (HRMS; Rocchetti e O’Callaghan, 2021). La recente applicazione della metabolomica mirata e non mirata al campo della scienza lattiero-casearia ha consentito di selezionare alcuni composti biomarcatori relativi alla salute degli animali, alla produzione, all’autenticazione e alle proprietà tecnologiche e funzionali del latte.
Il latte è un fluido biologico complesso prodotto dalle ghiandole mammarie dei mammiferi. Il sistema di alimentazione è riconosciuto come il principale fattore che influenza i processi metabolici dall’assunzione di nutrienti al rilascio delle ghiandole mammarie (Lamanna et al., 2011). Tra i fattori intrinseci più impattanti, è importante elencare le razze bovine e il periodo di allattamento (Xu et al., 2018, 2020), considerando la variazione stagionale, i sistemi di alimentazione, la gestione della mandria, l’origine geografica, le condizioni di salute della vacca da latte, la lavorazione, lo stoccaggio, e le condizioni tecnologiche rappresentano i fattori esterni più studiati, potenzialmente in grado di influenzare il metaboloma del latte. Per quanto riguarda i fattori esterni, l’effetto dei diversi sistemi di alimentazione sul profilo chimico del latte rappresenta uno degli argomenti di studio più recenti (O’Callaghan et al., 2018; Magan et al., 2019; Rocchetti et al., 2020, 2021 ; Bellassi et al., 2021; Yanibada et al., 2021). Inoltre, è noto che la combinazione di origine botanica e metodo di conservazione del foraggio modifica la composizione del latte, inclusi acidi grassi, vitamine liposolubili, composti N e acidi organici (O’Callaghan et al., 2016).
Nel lavoro scientifico pubblicato da Rocchetti et al., gli autori hanno valutato il potenziale della metabolomica non mirata, basata sulla cromatografia liquida ad altissima pressione (UHPLC) accoppiata con HRMS, per discriminare il profilo chimico dei campioni di latte sfuso secondo diverse strategie nutrizionali. La metabolomica del latte può essere utilizzata per catturare la maggior parte dei piccoli composti molecolari nel latte (come biofluido facilmente stoccabile e reperibile) e per identificare quelli che sono cambiati in modo significativo tra i diversi trattamenti dietetici. Questo approccio può discernere biomarcatori validi e percorsi metabolici discriminanti per comprendere meglio i complessi meccanismi che coinvolgono il sistema di alimentazione e la composizione del latte.
Questa sinossi è una sintesi dell’articolo scientifico pubblicato sul Journal of Dairy Science da Rocchetti et al. (2022) finalizzato a valutare il potenziale della metabolomica non mirata, basata sulla cromatografia liquida ad altissima pressione (UHPLC) accoppiata con HRMS (sfruttando l’Orbitrap come spettrometro di massa), per discriminare il profilo chimico dei campioni di latte di massa appartenenti a diverse strategie nutrizionali delle vacche da latte. La metabolomica del latte può essere utilizzata per catturare la maggior parte dei piccoli metaboliti nel latte (come biofluido facilmente prelevabile ed immagazzinabile) e per identificare i potenziali marcatori discriminanti di diversi regimi alimentari e strategie nutrizionali.
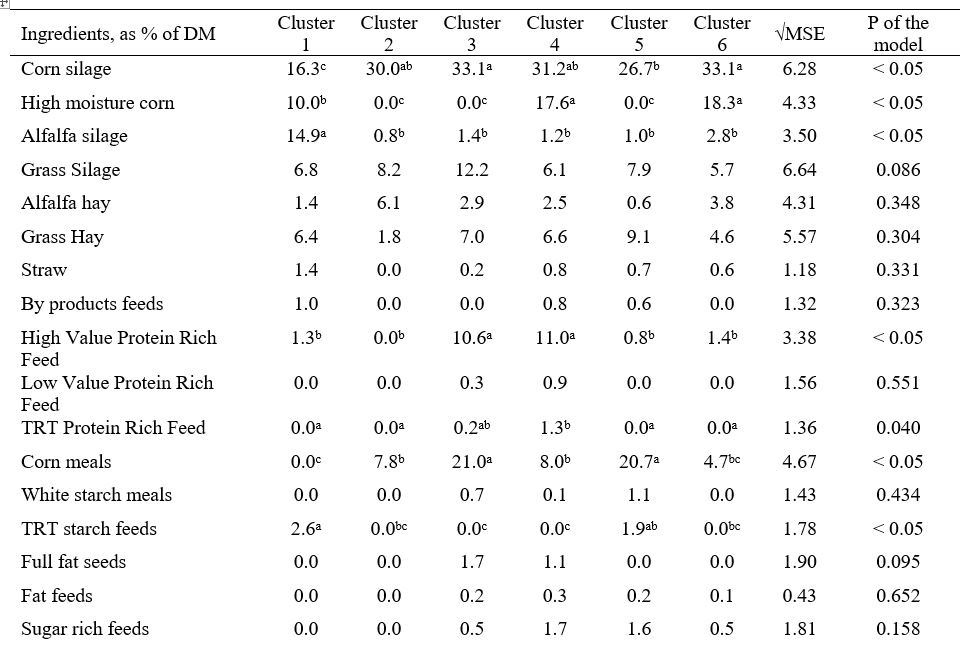
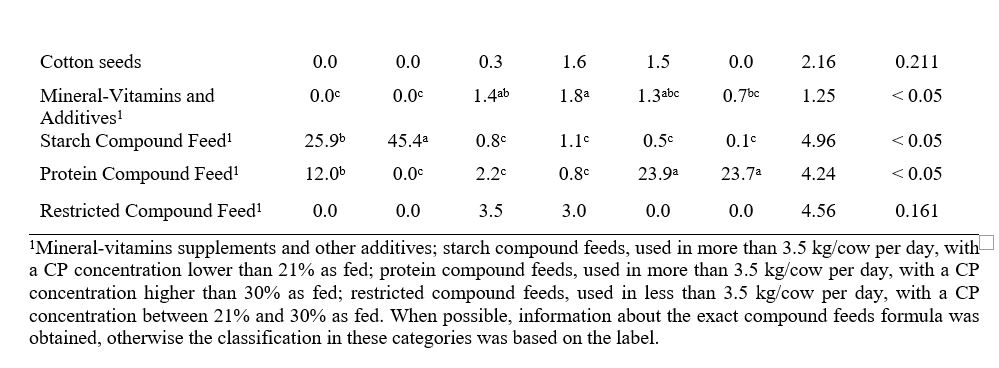
La scelta delle strategie nutrizionali da associare successivamente ad un determinato profilo chimico del latte è stata effettuata sulla base di un precedente lavoro pubblicato dallo stesso gruppo di ricerca (Gallo et al., 2022), dove è stato condotto uno studio osservazionale per verificare l’influenza di diverse strategie nutrizionali basate sull’insilato di mais sull’uso efficiente dei nutrienti, sul profilo di fermentazione fecale e sulla redditività in un coorte di aziende lattiero-casearie intensive (ossia un totale di 66 aziende lattiero-casearie commerciali di una specifica area d’Italia, cioè la Pianura Padana). Complessivamente, gli allevamenti avevano un’unica dieta per le vacche in lattazione, un’unica dieta per le vacche in asciutta e 2 diete per le manze (dallo svezzamento al primo parto), ad eccezione di 2 allevamenti che dividevano le vacche in lattazione in 2 gruppi, essendo precoci ( <150 DIM) e in tarda lattazione (>150 DIM). Un’analisi cluster gerarchica non supervisionata ha discriminato con successo le scelte alimentari dell’allevamento in 6 strategie nutrizionali principali, basate sull’uso elevato di (1) mais ad alta umidità (HMC) e insilato di legumi (cluster 1), (2) mangime concentrato (cluster 2), (3) farina di mais e soia (cluster 3), (4) HMC e farina di soia (cluster 4), (5) farina di mais e mangimi concentrati proteici (cluster 5) e (6) HMC e mangimi concentrati proteici (cluster 6). La Tabella 1 presenta le strategie (cluster) nutrizionali considerati in questo studio.
Tabella 1. Formulazione degli ingredienti considerando i diversi cluster basati sulla strategia nutrizionale (Gallo et al., 2022). I valori medi riportati con diverse lettere in apice all’interno di una stessa riga sono significativamente diversi (p < 0.05).
I campioni di latte di massa sono stati prelevati dalle 66 aziende e successivamente ne sono stati selezionati 36 in modo casuale per le successive analisi metabolomiche. Le analisi metabolomiche accoppiate all’utilizzo di analisi statistiche multivariate sono state effettuate in primis per valutare similarità e differenze nella composizione chimica dei latti e, successivamente, per valutare eventuali biomarcatori dei 6 principali cluster nutrizionali identificati da Gallo et al. (2022).
Lo screening basato sull’approccio metabolomico non mirato ha consentito l’identificazione putativa di 2.954 composti. Pertanto, per ridurre la complessità dei dati e fornire un dataset di composti da utilizzare per le seguenti analisi statistiche multivariate, sono stati sfruttati 3 database completi, vale a dire Bovine Metabolome Database, Phenol- Explorer e Mass Bank of North America. Seguendo questa strategia, è stata ottenuta l’identificazione accurata di 697 metaboliti del latte, confermando strutturalmente 88 metaboliti sulla base di analisi di spettrometria di massa tandem. In generale, tra i composti identificati, sono stati riscontrati soprattutto aminoacidi, peptidi, acidi carbossilici e derivati, composti legati all’alimentazione (come derivati dell’acido ippurico e metaboliti fenolici), derivati degli acidi nucleici (purine e pirimidine) e derivati delle piridine.
Il primo risultato che merita di essere evidenziato è la separazione iperspaziale fornita dallo score plot dell’analisi multivariata supervisionata OPLS-DA (Figura 1) e costruita considerando il profilo metabolomico dei diversi campioni di latte.
Figura 1. Score plot relativo all’analisi supervisionata OPLS-DA, sfruttata per raggruppare i campioni di latte in accordo al profilo chimico misurato per mezzo dell’analisi metabolomica untargeted.
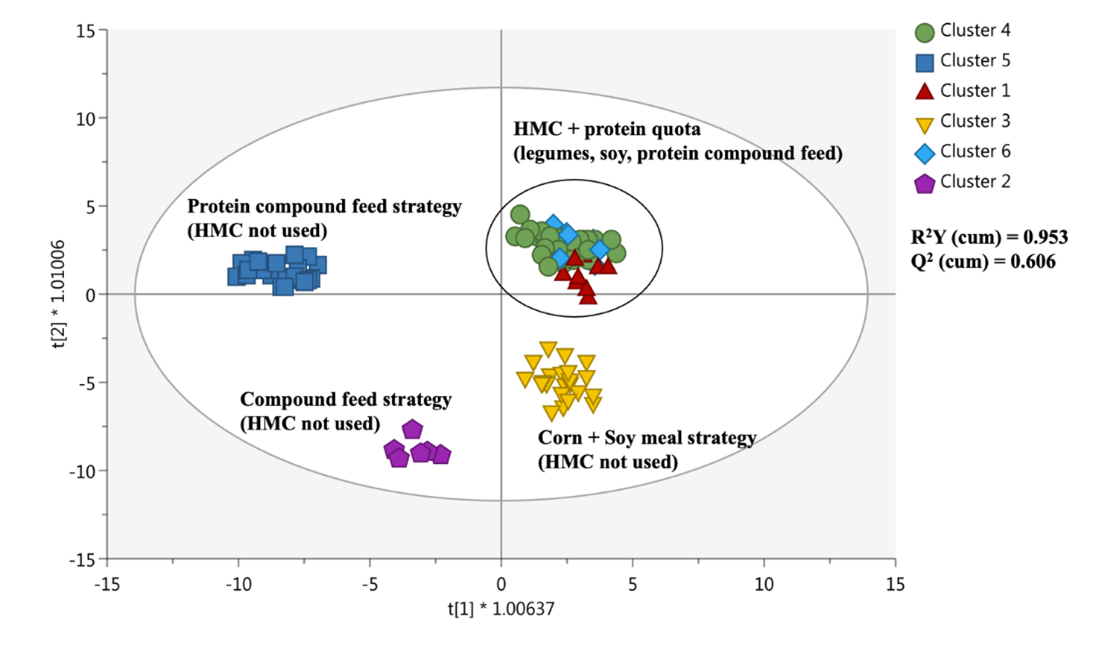
A questo proposito, è importante ricordare che la metabolomica untargeted (non mirata) consente di misurare un’ampia gamma di metaboliti presenti in un campione estratto senza nessuna conoscenza a priori del metaboloma e delle classi chimiche discriminanti. La separazione osservata in Figura 1 ha rivelato che i campioni di latte appartenenti ai cluster 1, 4 e 6 erano caratterizzati da un profilo chimico simile. Pertanto, è possibile ipotizzare un fattore discriminante comune che guida la separazione iperspaziale dei cluster 1 , 4 e 6. È interessante notare che il precedente studio osservazionale (Gallo et al., 2022) permetteva di identificare questo fattore trainante nell’utilizzo dell’HMC che caratterizzava esclusivamente (dal 10% fino al 18.3% della sostanza secca) la strategia nutrizionale dei cluster 1, 4 e 6 (Tabella 11). La potenziale correlazione tra l’uso di HMC nella razione e il profilo metabolomico del latte rappresenta un nuovo contributo al campo della scienza lattiero-casearia, poco esplorato nella letteratura scientifica.
Il passo successivo del lavoro è stato valutare l’effetto delle strategie di alimentazione basate sull’utilizzo di HMC sul profilo chimico del latte, estrapolando potenziali composti marcatori e vie metabolomiche discriminanti. Come riportato in Figura 2, l’analisi di arricchimento effettuata sui composti marcatori VIP del modello OPLS-DA ha consentito di osservare una rappresentanza significativa (p < 0.05) di metaboliti coinvolti nelle vie metaboliche dei derivati degli acidi nucleici (purina e pirimidine), AA (sia essenziali che non essenziali), peptidi e piridine (derivati piridossale).
Figura 2. Analisi statistica di arricchimento per evidenziare i pathway metabolici e le classi chimiche maggiormente discriminanti (p<0.05) il profilo chimico del latte in funzione delle strategie nutrizionali basate su HMC.
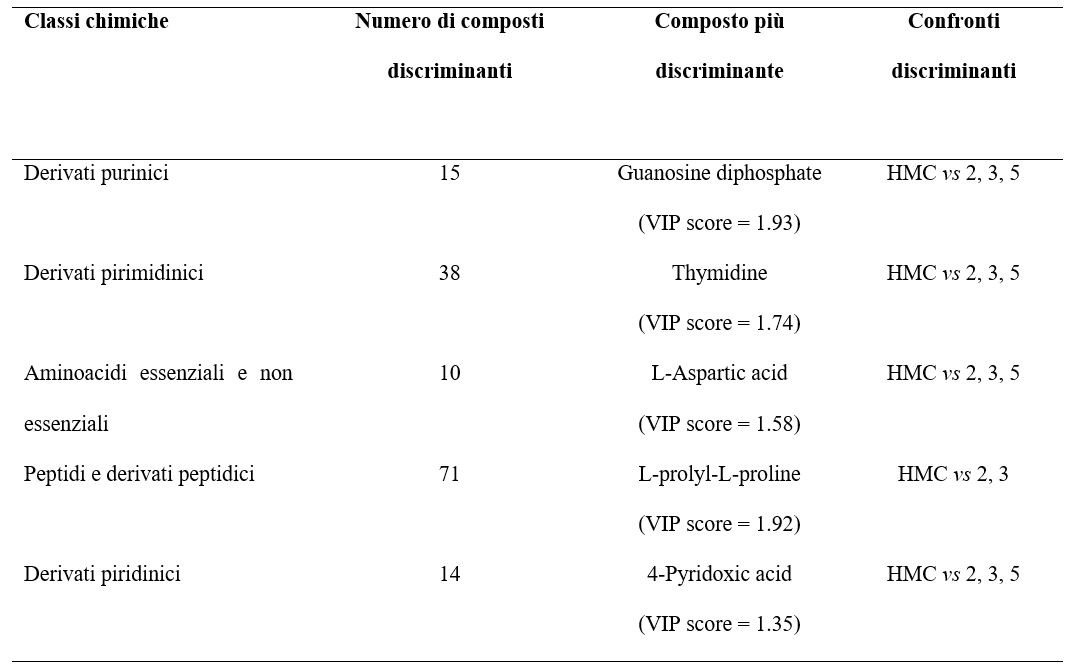
La maggior parte delle classi chimiche evidenziate tendevano a discriminare in modo significativo soprattutto il “cluster HMC” dagli altri 3 cluster (2, 3, e 5), come si evince dai valori tabulati nella Tabella 2.
Tabella 2. Classi chimiche e metaboliti maggiormente discriminanti il confronto HMC vs gli altri cluster associati a strategie nutrizionali prive di HMC. VIP score = punteggio discriminante.
Nel precedente lavoro pubblicato da Gallo et al. (2022), il livello di inclusione di HMC nelle diete aumentava l’efficienza del mangime (p < 0.05), la cinetica della produzione di gas (valore p <0.05), la proteina grezza, e la digeribilità dell’amido delle diete (p < 0.05). Inoltre, sono stati osservati livelli di produzione superiori a 33 kg/vacca al giorno quando si consideravano strategie nutrizionali a base di HMC (cluster 1, 4 e 6) nonostante le differenze in altri insilati o fonti di amido e proteine. Solitamente, una maggiore digeribilità dell’amido ruminale viene misurata quando le vacche da latte sono alimentate con HMC rispetto al mais secco (Firkins et al., 2001; Ferraretto et al., 2013). Ciò è probabilmente correlato alla rottura della matrice idrofobica dell’amido-proteina che circonda i granuli di amido durante l’insilamento (Gallo et al., 2014), che consente una maggiore fermentazione microbica e la digestione enzimatica dell’amido da parte dei ruminanti. Sebbene la digeribilità dell’amido dell’HMC possa essere alterata da diversi fattori (Gallo et al., 2022), è stato precedentemente descritto che l’HMC possiede un tasso di degradazione dell’amido ruminale maggiore rispetto alla farina di mais, essendo quest’ultima raccolta in uno stadio di maturità più avanzato (Masoero et al., 2011; Hoffman et al., 2012; Ferraretto et al., 2015), con potenziali effetti sulla degradabilità dell’amido e sulla sincronizzazione della fermentazione ruminale. Inoltre, precedenti lavori presenti in letteratura scientifica come quello di Eun et al. (2014), hanno scoperto che l’alimentazione di HMC in diete ad alto contenuto di foraggio ha aumentato la digeribilità di NDF e CP, la sintesi proteica microbica, insieme all’efficienza di utilizzo dei mangimi e dell’N, con una diminuzione del DMI rispetto al mais in fiocchi. Di conseguenza, le vacche alimentate con HMC possono avere una migliore efficienza energetica rispetto a quelle alimentate con mais in fiocchi di vapore. Tuttavia, non ci sono informazioni nella letteratura scientifica su come l’alimentazione con HMC influenzi il profilo metabolomico del latte e la composizione finale del latte e questo lavoro fornisce un’indicazione preliminare dei suoi effetti su un ampio pannello di diete per vacche da latte.
Nel loro insieme, i risultati basati sull’analisi metabolomica sembrano suggerire che l’inclusione di HMC nella formulazione dietetica ha avuto un grande effetto sul metabolismo dell’azoto, con particolare riferimento all’accumulo di acidi nucleici e ai loro prodotti di degradazione nel latte (Figura 3).
Figura 3. Pathway analisi che evidenzia i metabolismi del latte maggiormente discriminanti per i confronti HMC vs cluster 2, 3 e 5.
L’efficienza dell’azoto (N) nelle vacche da latte è bassa e l’ottimizzazione della dieta, con particolare attenzione alla % di N nella dieta sotto forma di proteine, aminoacidi ed urea, ha portato solo a piccoli miglioramenti nell’utilizzo di N nelle vacche da latte (Fujihara e Shem, 2011). Inoltre, l’importanza di altri composti contenenti N come gli acidi nucleici microbici (cioè, DNA e RNA) nella fisiologia nutrizionale dei ruminanti è stata finora scarsamente studiata, sebbene corrispondano a oltre il 20% dell’N microbico totale sintetizzato nel rumine.
Nel complesso, gli acidi nucleici di sintesi microbica fluiscono nell’intestino tenue dove vengono digeriti prima che avvenga il successivo assorbimento (Stentoft et al., 2015). Per quanto riguarda i biomarcatori specifici nel latte, l’analisi quantitativa dei derivati dell’acido nucleico nelle bovine da latte si è concentrata quasi esclusivamente sui derivati delle purine sia nell’urina che nel latte, dove i prodotti di degradazione delle purine acido urico e allantoina sono stati proposti come marcatori indiretti della sintesi microbica del rumine (Schager et al., 2003). In particolare, è stato ipotizzato che gli acidi nucleici che entrano nel duodeno siano quasi tutti di origine microbica. I nucleotidi purinici vengono degradati nell’intestino e quindi assorbiti. L’adenina e la guanina vengono catabolizzati ed escreti attraverso 4 possibili vie, vale a dire renale, ghiandola mammaria, riciclo attraverso la saliva e secrezione nell’intestino (Schager et al., 2003). I derivati delle purine vengono escreti principalmente come allantoina, ma anche come ipoxantina, xantina e acido urico. L‘allantoina e l’acido urico sono stati correlati all’elevata attività della xantina ossidasi nel sangue e nei tessuti, convertendo la xantina e l’ipoxantina in acido urico prima dell’escrezione. È interessante notare che, sebbene siano disponibili alcuni studi sull’estrazione dell’allantoina dal latte per stimare il flusso proteico microbico nel duodeno (Stentoft et al., 2015), i dati sui potenziali biomarcatori correlati ad altre purine e principalmente alle pirimidine sono attualmente molto limitati.
Nel lavoro pubblicato da Rocchetti et al. (2022), è stato riscontrato che il metabolismo delle purine possiede un effetto meno significativo sulle perturbazioni metabolomiche indotte dalle diete a base di HMC, mentre è possibile osservare un effetto maggiore del metabolismo delle pirimidine (Figura 3). Di conseguenza, nei campioni di latte correlati all’HMC sono stati trovati valori di accumulo più bassi sia dell’acido urico che dell’allantoina quando si considerava ogni possibile confronto (Tabella 3).

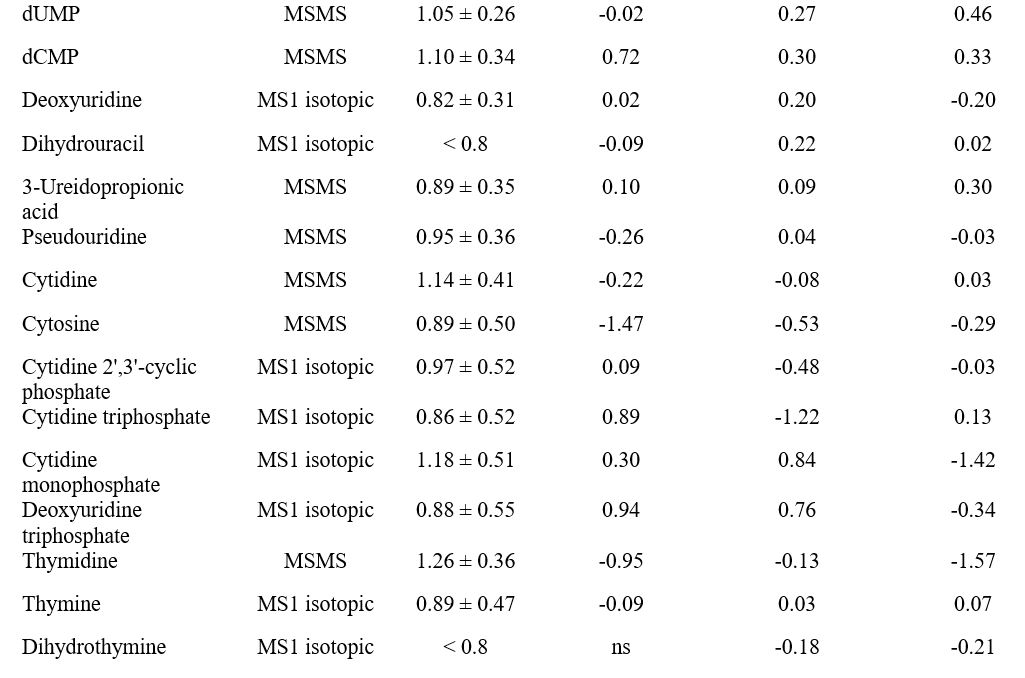
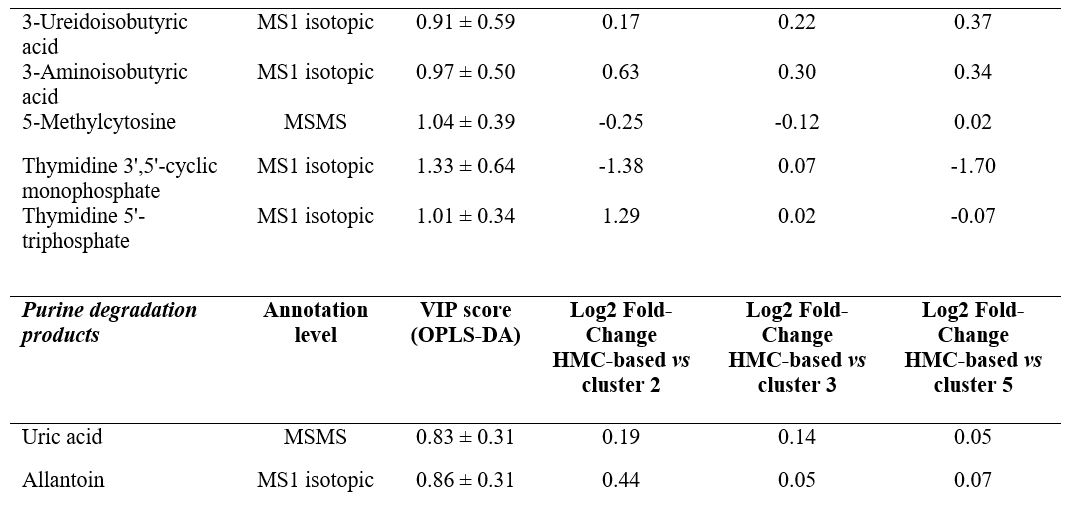
Nel complesso, l’ambiente ruminale ha una grande influenza sulla qualità del latte. Tuttavia, la correlazione tra i cambiamenti del profilo microbico e dei metaboliti del rumine insieme alla salute della mammella e ai profili metabolici del latte delle vacche da latte è ancora scarsa di conoscenza. I risultati di questo lavoro suggeriscono una potenziale rete condivisa nella fisiologia ruminale e nella produzione del latte considerando il significativo arricchimento non solo dei prodotti di degradazione delle pirimidine (come acido 3-ureidoisobutirrico, acido 3-aminoisobutirrico e acido 3-ureidopropionico), ma anche degli intermedi biosintetici pirimidinici (come L-aspartato, N-carbamoil-l-aspartato, derivati dell’acido orotico e nucleotidi; Tabella 3) necessari per la loro sintesi de novo (Löffler et al., 2015). Per quanto riguarda l’acido orotico, esso rappresenta una componente del latte non molto studiato ed è un indicatore del deficit metabolico delle vacche dell’enzima uridina monofosfato sintasi (Wehrmüller et al., 2008; Zaalberg et al., 2020). L’acido orotico è stato precedentemente descritto come un sottoprodotto della biosintesi proteica nel latte (Tiemeyer et al., 1984). Tuttavia, l’esatta funzione dell’acido orotico nel latte e il suo effetto sulla salute dei vitelli, la salute degli esseri umani che consumano latte o prodotti lattiero-caseari, le proprietà di produzione del latte e il suo potenziale come tratto indicatore, nonostante abbia attirato l’attenzione di diversi ricercatori in passato (Robinson, 1980; Tiemeyer et al., 1984; Saidi e Warthesen, 1989), sembra ancora in gran parte sconosciuto.
Nucleotidi, nucleosidi e nucleobasi appartengono alla frazione non proteica-azotata del latte, e si tratta di nutrienti dietetici semi-essenziali che caratterizzano il latte dei mammiferi ed esercitano molte funzioni fisiologiche. Tuttavia, l’efficienza della degradazione dipende in larga misura dall’attività enzimatica del rumine, che riflette le differenze nelle popolazioni microbiche tra animali e diete, e dal tasso di passaggio del rumine. A questo proposito, la quantità totale di DNA e RNA sintetizzata nel rumine dipende in gran parte dalla quantità di crescita batterica; pertanto, studi mirati che combinano il profilo metagenomico del microbiota ruminale e il profilo metabolomico di diversi biofluidi sembrano essere di grande interesse per l’industria casearia, considerando la necessità di misurare accuratamente l’apporto proteico microbico e di valutare meglio lo spostamento del microbiota ruminale verso il metabolismo delle pirimidine (come osservato in questo lavoro prendendo di mira il metaboloma del latte). Secondo la letteratura scientifica (Kanehisa et al., 2014), i metaboliti delle pirimidine vengono degradati durante il pre-assorbimento, nel sangue e nel tessuto epatico più o meno allo stesso modo dei metaboliti delle purine. Tuttavia, i dati sulle vie metaboliche delle pirimidine nei bovini sono attualmente molto limitati. È noto che i prodotti finali della degradazione delle pirimidine (come la β-alanina e l’acido β-aminoisobutirrico) possono agire come prodotti intermedi in altri metabolici legati all’azoto (come il metabolismo della β-alanina insieme alla degradazione di valina, leucina e isoleucina). Ciò potrebbe indicare che le vie di degradazione dei metaboliti delle pirimidine differiscono da quella dei metaboliti delle purine. In generale, non è noto quanto e quali tipi di metaboliti delle basi azotate vengono assorbiti dal lume intestinale e in che misura vengono metabolizzati attraverso il tessuto epatico (Stentoft et al., 2015). Per quanto ne sappiamo, la secrezione e il profilo completo dei metaboliti delle pirimidine nel latte non sono stati studiati nelle vacche da latte e la composizione finale della piridina nel latte rappresenta una complessa interazione che coinvolge i flussi splancnici totali dal rumine alla ghiandola mammaria. In un precedente lavoro di Stentoft et al. (2015), è stato studiato l’assorbimento e il metabolismo intermedio di purine e pirimidine nelle vacche da latte in lattazione. È interessante notare che gli autori hanno dimostrato che il metabolismo delle pirimidine differiva dal metabolismo delle purine nei tessuti splancnici, con una percentuale dell’afflusso totale di circa il 50%, suggerendo così che gli enzimi di degradazione delle pirimidine nel tessuto epatico non sono in grado di rimuovere gli intermedi e metaboliti pirimidinici che entrano dai tessuti periferici. Questa tendenza probabilmente rifletteva il fatto che livelli molto più elevati di nucleotidi pirimidinici e prodotti di degradazione entravano nel tessuto epatico e che l’efficacia del tessuto epatico e i requisiti o la tolleranza dell’organismo ai metaboliti erano diversi da quelli dei metaboliti delle purine. Le possibili conseguenze legate all’eccesso di tali metaboliti sono finora sconosciute; una possibile ipotesi, strettamente correlata ai risultati osservati nel lavoro di Rocchetti et al. (2022), potrebbe essere il loro riutilizzo nei tessuti periferici o l’escrezione finale nei biofluidi, come il latte. Pertanto, i risultati di questo lavoro sembrano supportare l’ipotesi che intermedi pirimidinici e prodotti di degradazione siano potenziali composti marcatori del complesso metabolismo dell’azoto microbico nelle vacche da latte e, per la prima volta, una correlazione indiretta tra dieta a base di HMC (descritta nella letteratura scientifica per aumentare la sintesi microbica ruminale) e derivati pirimidinici nel latte.
Concludendo, i ricercatori hanno utilizzato un approccio metabolomico per fornire nuove informazioni sulla potenziale correlazione esistente tra le diverse strategie nutrizionali delle vacche da latte e il profilo chimico del latte. Nel complesso, una combinazione di diversi approcci statistici multivariati ha permesso di identificare un potenziale effetto dell’HMC sul metabolismo delle pirimidine. L’accumulo di intermedi pirimidinici e dei loro prodotti di degradazione potrebbe essere considerato in futuro come potenziale marcatore di diete a base di HMC, note per essere caratterizzate da una maggiore fermentazione microbica e digestione enzimatica dell’amido da parte dei ruminanti. Inoltre, i nostri risultati metabolomici preliminari suggeriscono un complesso metabolismo dell’azoto microbico basato sui flussi splancnici dal rumine alla ghiandola mammaria nelle vacche da latte. Pertanto, ulteriori lavori, basati sulla valutazione completa di altri biofluidi (come rumine, plasma e urina) sembrano essere di grande interesse per chiarire meglio il ruolo biochimico e fisiologico dei metaboliti delle pirimidine.
La presente nota è una sintesi del seguente articolo scientifico pubblicato sul Journal of Dairy Science (Elsevier), dove è riportata tutta la letteratura citata: Rocchetti, G., Ghilardelli, F., Carboni, E., Atzori, A.S., Masoero, F., Gallo, A. (2022). Milk metabolome reveals pyrimidine and its degradation products as the discriminant markers of different corn silage-based nutritional strategies. Journal of Dairy Science, 105(11), 8650-8663.
Clicca qui per leggere l‘articolo integrale.
Autori:
Dr. Gabriele Rocchetti (Dipartimento di Scienze animali, della nutrizione e degli alimenti, Università Cattolica de Sacro Cuore), sotto la supervisione del “Gruppo Editoriale ASPA”: Giuseppe Conte, Alberto Stanislao Atzori, Fabio Correddu, Antonio Gallo, Antonio Natalello, Sara Pegolo, Manuel Scerra.





Scrivi un commento
Devi accedere, per commentare.